|
 |
|
|
Chapter 9
Soil Aeration (contd.)The Harmful Effect of Grass
The harmful effect of grass on fruit trees varies with the species and with the period in the life of the tree when the grass is planted. Young trees are more adversely affected than fully developed individuals, which contain large quantities of reserves in the wood. Deciduous species suffer more than evergreens. These facts suggest that the harmful effect of grass is a consequence of starvation.
The effect of grass on young trees was first studied. The custard apple was the most sensitive. The trees were killed in 1916 within the first two years after the grass was planted. Next in order of susceptibility were the loquat (all died before the end of 1919), the plum, the lime, and the peach. The litchi and the mango just managed to maintain themselves. The guava was by far the least affected, the trees under grass being almost half the height of those under clean cultivation.
Grass not only reduces the amount of new growth but affects the leaves, branches, old wood, and fruit as well as the root system. The results relating to the above ground portion of the trees closely follow those described by the Woburn investigators. Compared with the foliage produced under clean cultivation, the leaves from the trees under grass appear later, are smaller and yellower and fall prematurely. The internodes are short. The bark of the twigs is light coloured, dull, and unhealthy and quite different from that of healthy trees. The bark of the old wood has a similar appearance and attracts lichens and algae to a much greater extent than that of the cultivated trees. The trees under grass flower late and sparingly. The fruit is small, tough, very highly coloured, and ripens earlier than the normal.
The effect of grass on the root system is equally striking. Except in the guava, the effect of grass on the superficial system is to restrict the amount of root development, to force the roots below the grass, and to reduce the number of active roots during the monsoon. The guava is an exception. The surface system is well developed, the roots are not driven downwards by the grass while active rootless are readily formed in the upper 4 inches of soil soon after the rains begin, very much as in the cultivated trees. In August 1922, when the ground water had risen to its highest point, the absorbing roots of the guava were found in the surface film of soil, and also above the surface among the stems of the grass. The grass carpet therefore acts as an asphyxiating agency in all these species, the guava excepted.
The grass covering has no appreciable effect either on the development or on the activity of the deep roots. This portion of the root system was explored during the hot weather of 1921 in the case of the guava (Plate VI, Fig. 3), mango, and litchi and results were obtained very similar to those in the corresponding cultivated trees.
Grass not only affects the roots underneath but also the development of those of the neighbouring trees under cultivation. Such roots either turn away from the grass, as in the custard apple, or else turn sharply downwards before they reach it.
A number of conclusions can be drawn from these root exposures. The custard apple, loquat, peach, and lime are unable to maintain their surface root systems under grass, but behave normally as regards the deep root system. Only the guava is able to get its roots above those of the grass during the rains.
The study of the harmful effect of grass on established trees also yielded interesting results. In this case the trees carried ample reserves in the wood and, as might be expected, the damage was less spectacular than in the case of young trees with little or no reserves. The order of susceptibility to grass, however, was very much the same in the two cases. When the fully-grown trees were first put under grass in August 1921, the grass at first grew poorly in tufts with bare ground between. Even this imperfect covering soon affected the custard apples, loquats, peaches, and litchis. By the rains of 1922 the grass became continuous; the effect on the trees was then much more marked.
In the plum interesting changes occurred. In July 1922, less than a year after planting the grass, the new shoots showed arrested growth and the foliage was attacked by leaf-destroying insects, which, however, ignored the leaves of the neighbouring cultivated plot. If the insects were the real cause of the trouble, it is difficult to see why the infection did not spread beyond the trees under grass. In January 1923 the average length of the new wood in these trees was 1 foot 5 inches compared with 3 feet 7 inches in the controls. The twigs were dull and purplish, the internodes were short (Plate V, Fig. 5). In February 1923 flowering was restricted and in April only tufts of leaves were formed at the ends of the branches instead of new shoots (Plate V, Fig. 8). Early in 1924, when I left Pusa and had to discontinue the work, a great deal of die-back was taking place.
Very similar results were obtained in all the species except the mango, which resisted grass better than any of the others. No definite effect was observed in this species till June 1923, when the foliage became distinctly lighter than that of the cultivated trees. The general results brought about by grass in all these cases suggested that the trees were slowly dying from starvation.
A year after the grass was planted and the grass effect was becoming marked, the root system of these established trees was examined. In August 1922 the plums, peaches, custard apples, mangoes, litchis, and loquats under grass were found to have produced very few active rootless in the upper foot of soil compared with the controls. In the case of the custard apples and the loquats, which suffered most from grass, there was a marked tendency for the new roots to grow downwards and away from the grass. No differences were observed in the dormancy or activity of the deep root system as compared with the controls. The deep roots behaved exactly like those under clean cultivation.
FIG. 4. The effect of burrowing rats on the growth of the plum under grass (June 21st, 1923)
During these examinations two instances of the striking effect of increased aeration on root development were observed. In July 1923 burrowing rats took up their quarters under one of the limes and one of the loquats, in each case on the southern side. Shortly afterwards the leaves just above the rat holes became very much darker in colour than the rest. Examination of the soil immediately round the burrows showed a copious development of new active rootless, far greater even than in the surface soil of the cultivated plot. The extra aeration had a wonderfully stimulating effect on the development of active roots, even under grass. The appearance of the leaves suggested an application of nitrogenous manure. Similar observations were made in the case of the plum (Fig. 4). Here the burrows caused a dying tree to produce new growth.The Effect of Aeration Trenches on Young Trees under Grass
The effect of aeration trenches in modifying the influence of grass suggests that one of the factors at work is soil asphyxiation. In the case of the custard apple and the lime the aeration trenches had no effect; all the trees died. The death of the plums was delayed by the aeration trenches. The loquats, litchis, and mangoes benefited considerably. In the guavas the trees provided with aeration trenches were indistinguishable from those under grass. The general results are shown in Table 8, in which the measurements of a hundred fully-developed leaves, made in March 1921, are recorded.
Table 8
The reduction in leaf size under grassxx Grass Grass with aeration trenches Cultivated xx cm. cm. cm. Plum 3.2 x 1.1 4.6 x 1.7 7.1 x 2.9 Peach 7.1 x 1.8 8.2 x 2.3 11.4 x 3.1 8.1.x 3.2 10.6 x 4.4 11.3 x 4.4 Mango 11.2 x 2.9 13.7 x 3.8 20.9 x 5.5 Litchi 8.9 x 2.4 11.5 x 3.4 12.2 x 3.5 Lime 3.8 x 1.6 5.2 x 2.1 6.4 x 3.4 Loquat Trees dead 16.4 x 4.6 22.1 x 5.9
At the end of 1920 the roots were exposed to a depth of 2 feet in order to ascertain the effect of the extra aeration on the development of the superficial system. The results were interesting. In all cases the superficial roots were much larger and better developed than those under grass, except in the guava where no differences in size could be detected. The roots were attracted by the trenches, often branching considerably in the soil at the side of the trenches themselves. The aeration trenches are made use of only during the monsoon phase. After the break of the rains, new active roots are always found in or near the trenches first, after which a certain amount of development takes place under the grass.
The deep root system of the trees provided with aeration trenches behaved exactly like the controls.The Results Obtained
The general results obtained with clean cultivation, grass, and grass with aeration trenches are shown in Plate IV, in which representative trees from the various plots have been drawn to scale. The drawings give a good idea of the main results of the experiment, namely: (1) the extremely deleterious effect of grass on young trees; (2) the less harmful effect of the same treatment on mature trees; (3) the partial recovery which sometimes takes place from the aeration trenches; and (4) the exceptional nature of the results with the guava, where the trees are able to grow under grass, but with reduced vigour, and where the aeration trenches have had little or no effect.
As would be expected from these results even a temporary removal of the grass cover has a profound effect. Whenever the roots of a tree under grass are exposed (for which purpose the grass has to be removed for a few days) there is an immediate increase in growth, accompanied by the formation of larger and darker-coloured leaves. The effect is clearly visible in the foliage above the excavation for as long as two years, but the rest of the tree is not affected.The Cause of the Harmful Effect of Grass
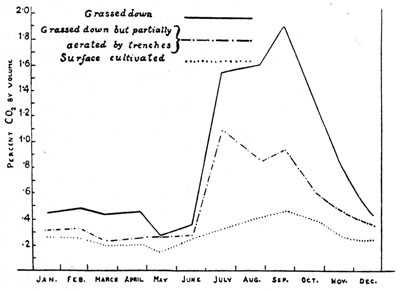
The examination of the root system of these eight species suggested that the first step in working out the cause of the harmful effect of grass would be to make a periodical examination of the soil gases. Determinations of the amount of CO2 in the soil-air at a depth of 9 to 12 inches were carried out during 1919 under grass, under grass with aeration trenches, and under cultivated soil. About 10 litres of air were drawn out of the soil at each determination and passed through standard baryta which was afterwards titrated in the ordinary way. The 1919 results are given in Table 9 and are set out graphically in Fig. 5.
Table 9
Percentage by volume of carbon dioxide in the soil-gas, under grass and clean cultivation, Pusa, 1919*Date and month when soil-gas was aspirated and analysed Plot no. 1 grassed Plot no. 2 grassed, but partially aerated by trenches Plot no. 3 surface cultivated Rainfall in inches since January 1st, 1919 January 13, 14 and 17 0.444 0.312 0.269 Nil February 20 and 21 0.472 0.320 0.253 1.30 March 21 and 22 0.427 0.223 0.197 1.33 April 23 and 24 0.454 0.262 0.203 2.69 May 16 and 17 0.271 0.257 0.133 3.26 June 17 and 18 0.341 0.274 0.249 4.53 July 17 and 18 1.540 1.090 0.304 14.61 August 25 and 26 1.590 0.836 0.401 23.29 September 19 and 20 1.908 0.931 0.450 30.67 October 21 and 22 1.297 0.602 0.365 32.90 November 14 and 15 0.853 0.456 0.261 32.90 December 22 and 23 0.398 0.327 0.219 32.92 *The determinations were carried out by Mr Jatindra Nath Mukerjee of the Chemical Section, Pusa.
The results of 1920 and 1921 confirm these figures in all respects. Table 9 shows that during the monsoon the volume of carbon dioxide in the pore spaces under grass is increased about fivefold in comparison with the soil-air of cultivated land. As this gas is far more soluble in water than oxygen, the amounts of carbon dioxide actually dissolved in the water-films in which the root-hairs work would be much higher than the figures in the table suggest.
The production of large amounts of carbon dioxide in the soil-air during the rains would also affect the formation of humus, nitrification, and the mycorrhizal relationship, all of which depend on adequate aeration. Considerable progress was made in the investigation of the supply of combined nitrogen. At all periods of the year, except at the break of the rains, the amount of nitric nitrogen in the upper 18 inches of soil under grass varied from 10 to 20 per cent. of that met with in the cultivated plots. When the shortage of nitrogen in the case of the guava was made up by means of sulphate of ammonia during the rains of 1923, the trees under grass at once responded and produced fruit and foliage hardly distinguishable in size from the controls.
In the case of the litchi and loquat, the roots of which are unable to aerate themselves in the rains by forcing their way through the grass to the surface, heavy applications of combined nitrogen improved the growth, but a distinctly harmful effect remained -- the manured trees as regards size and colour of the leaves, time of flowering, and production of new shoots occupying an intermediate position between the unmanured trees under grass and those under clean cultivation. These results are very similar to those obtained with apples at Cornell. At both places grass led to the disappearance of nitrates in the soil and restricted root development. The effect was only partially removed by the addition of nitrate of soda.; In the guava, however, combined nitrogen removes the harmful effect because the roots of this tree are able to obtain all the oxygen they need. The guava, therefore, suffers from only one of the factors resulting from a grass carpet -- lack of nitrate. The litchi and the loquat suffer from another factor as well -- lack of oxygen.
FIG. 5. Carbon dioxide in soil atmosphere, Pusa, 1919.Forest Trees and Grass
Although the grass carpet acts as an asphyxiating agent to the roots of all the fruit trees investigated except the guava, the ordinary Indian forest trees thrive under grass. Between the years 1921 and 1923 the relation between the grass carpet and the roots of the following fifteen forest trees was investigated (Table 10). All thrive remarkably well under grass and show none of the harmful effects exhibited by fruit trees.
Most of the forest trees in the plains of India flower and come into new leaf in the hot season and then proceed to form new shoots. After the early rains a distinct change is visible in the size, colour, and appearance of the foliage. The leaves become darker and more glossy; the story told by the young shoots of the custard apple is repeated.
Table 10
Forest trees under grass in the Botanical Area, PusaSpecies Time of flowering Time of leaf-fall Polyalthia longifolia Benth. & Hook, f. February-April April Melia Azadirachta L. March-May March Ficus bengalensis L. April-May March Ficus religiosa L. April-May December Ficus infectoria Roxb. February-May December-January Millingtonia hortensis Linn., f. November-December March Butea frondosa Roxb. March February Phyllanthus Emblica L. March-May February April-June March-April Tectonia grandis Linn., f. July-August February-March Thespesia populnea Corr. Throughout the year but chiefly in the cold season April Pterospermum acerifolium Willd. March-June January-Feburary Wrightia tomentosa Roem. & Schult. April-May January-February Lagerstroemia Flos-Regina Retz May December-January Dalbergia Sissoo Roxb. March December-January
Examination of the superficial root systems of the fifteen species during the rains of 1922 and 1923 yielded remarkably uniform results. All the trees produced abundant, normally developed active rootless in the upper 2 or 3 inches of soil and also on the surface; they therefore compete successfully with grass both for oxygen and nitrates. The large superficial roots were also well developed and compared favourably with the corresponding root system of fruit trees under clean cultivation. The grass carpet had apparently no harmful effect on the root system near the surface.
Between the hot weather of 1921 and the early months of 1924 the complete root systems of these fifteen species were investigated. In all cases the large surface roots gave off thin branches which grew vertically downwards to the cold-season level of the ground water. Root activity in all cases was practically confined in the hot season to the deep moist layers of sand between 10 and 20 feet below the surface, the roots always making the fullest use of the tunnels of Termites and other burrowing insects for passing easily through clay layers from one zone of sandy soil to the next below. Cavities in the soil were always fully used for root development. Soon after the rains the dormant surface roots burst into activity. As the ground water rose the deep root system became dormant; in August the active surface roots always showed marked aerotropism. The formation of nitrates which takes place about the time the cold-season crops are sown was followed by a definite burst of renewed root activity in the surface soil, followed by the production of new shoots and leaves. As the ground water falls in the autumn and the soil draws in oxygen, the formation of active roots follows the descending water-table exactly as has been described in the case of the guava.
The facts of root distribution and periodicity in root activity in forest trees explain why these trees do so well under grass and are able to vanquish it if allowed free competition. The chief weapons which enable forest trees to oust grasses and herbs from the habitat are the following:
- The deep root system admits of growth during the dry season when the grass is dormant, thereby enabling the trees to utilize moisture and food materials in the soil down to at least 20 feet. This markedly extends the period of assimilation.
- The habit of trees is a great advantage in the struggle for light.
- The active roots of the surface system are resistant to poor soil aeration, and are able to reach the surface and compete successfully with the grass for oxygen and for minerals.
The character which distinguishes forest trees from fruit trees is the power possessed by the surface roots of the former to avoid the consequences of poor soil aeration by forcing their way through a grass carpet in active growth to the air and to obtain oxygen as well as a share of the nitrates in the surface soil. The surface roots of most fruit trees are very susceptible to carbon dioxide and try to avoid it by growing downwards. The trees are therefore deprived of oxygen and of combined nitrogen during the rains, and slowly starve. The guava is an exception among fruit trees. Here the active roots reach the surface in the rains and the trees are able to maintain themselves. This explains why the pastures of Grenada and St. Vincent in the West Indies are so rapidly invaded and destroyed by the wild guava. The hedgerows and pastures of Great Britain if left to themselves behave in a similar way. The hedgerows soon begin to invade the fields. Young trees make their appearance; grass areas become woodland. The transformation, however, is much slower in Great Britain than in the tropics.
These studies on the root development of tropical forest trees throw a good deal of light on the soil aeration factor and the part the plant can play in such investigations. The movement of the ground water affects soil aeration directly. The two periods -- the beginning and end of the rainy season -- when the surface soil contains abundant air and ample moisture and when the temperature is favourable for nitrification, correspond exactly with times when nitrates accumulate and when growth is at its maximum. When soil aeration is interfered with during the rains by two factors, (1) the rise of the ground water, and (2) the formation of colloids in the surface soil, the plant roots respond by growing to the surface. Root development, therefore, is an important instrument in such an investigation when examined throughout the year.
The root development of trees influences the maintenance of soil fertility in the plains of India and indeed in many other regions. The dead roots provide the deeper layers of soil with organic matter and an almost perfect drainage and aerating system. The living roots comb the upper 20 feet of soil for such minerals as phosphates and potash which are used in the green leaves. These leaves in due course are converted into humus and help to enrich the surface soil. This explains why the soils of North Bihar, although very low in total and available phosphates, are so exceedingly fertile and yield heavy crops without any addition of mineral manures. The figures given by the analysis of the surface soil must be repeated in the lower layers and should be interpreted not in terms of the upper 9 inches but of the upper 20 feet.
The tree is the most efficient agent available for making use of the minerals in the soil. It can grow almost anywhere, it will vanquish most of the other forms of vegetation, and it will leave the soil in a highly fertile condition. It follows therefore that the trees and shrubs of the hedgerows, parks, and woodlands of countries like Great Britain must continue to be used for the maintenance of soil fertility. In Saxon times most of our best land was under forest. The fertility stored in the soil made the gradual clearing of this woodland worth while. In the future, when agriculture comes into its own and when it is no longer regarded solely as an industry, it may be desirable to embark on long term rotations in which woods and park-land are turned into arable, and worn-out arable back into woodland or into mixed grass and trees. In this way the root system of the tree can be used to restore soil fertility.The Aeration of the Sub-soil
One of the universal methods of improving aeration is subsoiling. The methods adopted vary greatly according to the factor which has interfered with aeration and the means available for improving the air-supply.
In temperate regions the chief factor which cuts off the sub-soil from the atmosphere is shortage of humus aggravated by impermeable pans (produced by the plough and by the soil particles themselves) or a permanent grass carpet accompanied by the constant treading of animals. The result in all cases is the same -- the supply of air to the sub-soil is reduced.
In loamy soils plough-pans develop very rapidly if the content of organic matter falls off and the earthworm population declines. A well-defined zone of close and sticky soil is formed just under the plough sole which holds up water, thereby partly asphyxiating the sub-soil below and water-logging the soil above.
In sandy soils as well as in silts, pans are formed with the greatest ease from the running together of the particles, particularly when artificials take the place of farm-yard manure and the temporary ley is not properly utilized. One of the most interesting cases of pan formation that I have observed in Great Britain was on the permanent manurial plots of the Woburn Experiment Station, where an attempt to grow cereals year after year on the greensand by means of artificial manures has been followed by complete failure of the crop. The soil has gone on permanent strike. The destruction of the earthworm population by the regular application of chemicals had deprived the land of its natural aerating agencies. Failure to renew the organic matter by a suitable rotation had resulted in a soil devoid of even a trace of tilth. About 9 inches below the surface, a definite pan (made up of sand particles loosely cemented together) occurred, which had so altered the aeration of the sub-soil that the whole of these experimental plots were covered with a dense growth of mares' tail (Equisetum arvense L.), a perennial weed which always indicates a badly aerated sub-soil. Nature as usual had summed up the position in her own inimitable fashion. There was no need of tabulated yields, analyses, curves, and statistics to explain the consequences of improper methods of agriculture.
The conventional method of dealing with arable pans in this country is by means of some sub-soiling implement which breaks them up and restores aeration. This should be accompanied whenever possible by heavy dressings of farm-yard manure, so that the tilth can be improved and the earthworm population restored. Some deep-rooted crop like lucerne, or even a temporary fey, should be called in to complete the cure. Sub-soiling heavy land under grass is proving even more advantageous than on arable areas. This leads, as we have seen, to humus formation under the turf and to an increase in the stock-carrying capacity of the land.
In the East the ventilation of the sub-soil is perhaps even more important than in the West. In India, for example, one of the common consequences of the monsoon rainfall and of flooding the surface with irrigation water is pan formation on a colossal scale due to the formation of soil colloids -- the whole of the surface soil tends to become a pan. This has to be broken up. The cultivators of the Orient set about this task in a very interesting way. Whenever they can use the roots of a leguminous crop as a sub-soiler they invariably employ this machine. It has the merit of costing nothing, of yielding essential food and fodder, and of suiting the small field. In the Indo-Gangetic plain the universal sub-soiler is the pigeon pea, the roots of which not only break up soil pans with ease but also add organic matter at the same time. On the Western frontier the sub-soiling of the dense loess soils is always done by the roots of a lucerne crop. On the black cotton soils of Peninsular India where the monsoon rainfall converts the whole of the surface soil into a vast colloidal pan, the agricultural situation is saved by the succeeding hot season which dries out this pan and reduces its volume to such an extent that a multitude of deep fissures occur right down to the sub-soil. The black soils of India plough and sub-soil themselves. The moist winds, which precede the south-west monsoon in May and early June, replace some of the lost moisture; the heavy clods break down and when the early rains arrive a magnificent tilth can be prepared for the cotton crop. The sub-soiling in this case is done by Nature; the cultivators merely give a subsequent cultivation and then sow the crop.Bibliography
Clements, F. E. Aeration and Air Content: the Role of Oxygen in Root Activity, Publication No. 315, Carnegie Institution of Washington, 1921.
Howard, A. Crop Production in India: A Critical Survey of its Problems, Oxford University Press, 1924.
-- 'The Effect of Grass on Trees', Proc. Royal Soc., Series B, xcvii, 1925, p. 284.
Lyon, T. L., Heinicke, A. J., and Wilson, D. D. The Relation of Soil Moisture and Nitrates to the Effects of Sod on Apple Trees, Memoir 63, Cornell Agricultural Expt. Station, 1923.
The Duke of Bedford, and Pickering, S. U. Science and Fruit-Growing, London, 1919.
Weaver, J. E., Jean, F. C, and Crist, J. W. Development and Activities of Crop Plants, Publication No. 316, Carnegie Institution of Washington, 1922.
Next: 10. Some Diseases of the Soil
Back to Contents
To Albert Howard review and index
Back to Small Farms Library index
Community development | Rural development
City farms | Organic gardening | Composting | Small farms | Biofuel | Solar box cookers
Trees, soil and water | Seeds of the world | Appropriate technology | Project vehicles
Home | What people are saying about us | About Handmade Projects
Projects | Internet | Schools projects | Sitemap | Site Search | Donations |